
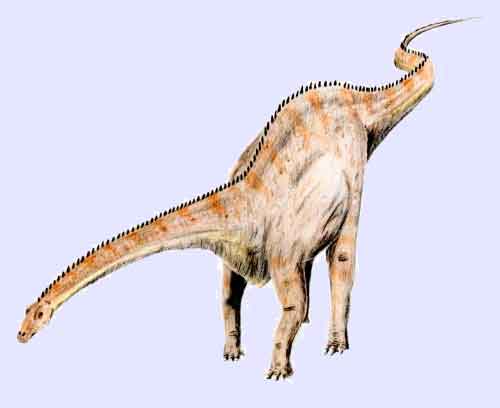
Illustration of Diplodocus carnegiei (*)
Diplodocus (pronounced /dɪˈplɒdəkəs/,[1][2] /daɪˈplɒdəkəs/,[2] or /ˌdɪploʊˈdoʊkəs/[1]) is a genus of diplodocid sauropod dinosaur whose fossils were first discovered in 1877 by S. W. Williston. The generic name, coined by Othniel Charles Marsh in 1878, is a Neo-Latin term derived from Greek διπλόος (diploos) "double" and δοκός (dokos) "beam",[1] in reference to its double-beamed chevron bones located in the underside of the tail. These bones were initially believed to be unique to Diplodocus; however, they have since then been discovered in other members of the diplodocid family and in non-diplodocid sauropods such as Mamenchisaurus.
It lived in what is now western North America at the end of the Jurassic Period. Diplodocus is one of the more common dinosaur fossils found in the Upper Morrison Formation, a sequence of shallow marine and alluvial sediments deposited about 150 to 147 million years ago, in what is now termed the Kimmeridgian and Tithonian stages. The Morrison Formation records an environment and time dominated by gigantic sauropod dinosaurs such as Camarasaurus, Barosaurus, Apatosaurus and Brachiosaurus.[3]
Diplodocus is among the most easily identifiable dinosaurs, with its classic dinosaur shape, long neck and tail and four sturdy legs. For many years, it was the longest dinosaur known. Its great size may have been a deterrent to the predators Allosaurus and Ceratosaurus: their remains have been found in the same strata, which suggests they coexisted with Diplodocus.
Description
One of the best-known sauropods, Diplodocus was a very large long-necked quadrupedal animal, with a long, whip-like tail. Its forelimbs were slightly shorter than its hind limbs, resulting in a largely horizontal posture. The long-necked, long-tailed animal with four sturdy legs has been mechanically compared with a suspension bridge.[4] In fact, Diplodocus is the longest dinosaur known from a complete skeleton.[4] The partial remains of D. hallorum have increased the estimated length, though not as much as previously thought; when first described in 1991, discoverer David Gillete calculated it may have been up to 54 m (177.05 ft) long, making it the longest known dinosaur (excluding those known from especially poor remains, such as Amphicoelias). Some weight estimates ranged as high as 113 (rather only 50) tonnes (125 US short tons). This review was based on recent findings that show that the giant tail vertebrae were actually placed further forward on the tail than Gillete originally calculated. The study shows that the complete Diplodocus skeleton at the Carnegie Museum of Natural History in Pittsburgh, Pennsylvania on which estimates of Seismosaurus were based had its 13th tail vertebra come from another dinosaur, throwing size estimates for Seismosaurus off by up to 30%. While dinosaurs such as Supersaurus were probably longer, fossil remains of these animals are only fragmentary.[5]
The skull of Diplodocus was very small, compared with the size of the animal, which could reach up to 35 m (115 ft),[6] of which over 6 m (20 ft) was neck.[7] Diplodocus had small, 'peg'-like teeth that pointed forward and were only present in the anterior sections of the jaws.[8] Its braincase was small. The neck was composed of at least fifteen vertebrae and is now believed to have been generally held parallel to the ground and unable to have been elevated much past horizontal.[9] Modern mass estimates have tended to be in the 10 to 16 tonne (11–17.6 ton) range: 10 tonnes (11 tons);[10] 11.5 tonnes (12.7 tons);[11] 12.7 tonnes (14 tons);[12] and 16 tonnes (17.6 tons).[13]
Diplodocus had an extremely long tail, composed of about 80 caudal vertebrae,[14] which is almost double the number some of the earlier sauropods had in their tails (such as Shunosaurus with 43), and far more than contemporaneous macronarians had (such as Camarasaurus with 53). There has been speculation as to whether it may have had a defensive[15] or noisemaking function.[16] The tail may have served as a counterbalance for the neck. The middle part of the tail had 'double beams' (oddly-shaped bones on the underside, which gave Diplodocus its name). They may have provided support for the vertebrae, or perhaps prevented the blood vessels from being crushed if the animal's heavy tail pressed against the ground. These 'double beams' are also seen in some related dinosaurs.
Like other sauropods, the manus (front "feet") of Diplodocus were highly modified, with the finger and hand bones arranged into a vertical column, horseshoe-shaped in cross section. Diplodocus lacked claws on all but one digit of the front limb, and this claw was unusually large relative to other sauropods, flattened from side to side, and detached from the bones of the hand. The function of this unusually specialized claw is unknown.[17]




Diplodocus , Diplodocus , Diplodocus , Diplodocus ,
Discovery and species
Several species of Diplodocus were described between 1878 and 1924. The first skeleton was found at Como Bluff, Wyoming by Benjamin Mudge and Samuel Wendell Williston in 1878, and was named Diplodocus longus ('long double-beam'), by paleontologist Othniel Charles Marsh in 1878.[18] Diplodocus remains have since been found in the Morrison Formation of the western U.S. States of Colorado, Utah, Montana and Wyoming. Fossils of this animal are common, except for the skull, which is often missing from otherwise complete skeletons. Although not the type species, D. carnegiei is the most completely known and most famous due to the large number of casts of its skeleton in museums around the world.
The two Morrison Formation sauropod genera Diplodocus and Barosaurus had very similar limb bones. In the past, many isolated limb bones were automatically attributed to Diplodocus but may, in fact, have belonged to Barosaurus.[19]
Valid species
Seismosaurus redirects here. For the fictional mecha based on this dinosaur, see Seismosaurus (Zoids)
* D. longus, the type species, is known from two skulls and a caudal series from the Morrison Formation of Colorado and Utah.[6]
* D. carnegiei, named after Andrew Carnegie, is the best known, mainly due to a near-complete skeleton collected by Jacob Wortman, of the Carnegie Museum of Natural History in Pittsburgh, Pennsylvania and described and named by John Bell Hatcher in 1901.
* D. hayi, known from a partial skeleton discovered by William H. Utterback in 1902 near Sheridan, Wyoming, was described in 1924.[20]
* D. hallorum, first described in 1991 by Gillette as Seismosaurus halli from a partial skeleton comprising vertebrae, pelvis and ribs. George Olshevsky later attempted to emend the name as S. hallorum, citing incorrect grammar on the part of the original authors, a recommendation that has been followed by others, including Carpenter (2006).[21] In 2004, a presentation at the annual conference of the Geological Society of America made a case for Seismosaurus being a junior synonym of Diplodocus.[22] This was followed by a much more detailed publication in 2006, which not only renamed the species Diplodocus hallorum, but also speculated that it could prove to be the same as D. longus.[23] The position that D. hallorum should be regarded as a specimen of D. longus was also taken by the authors of a redescription of Supersaurus, refuting a previous hypothesis that Seismosaurus and Supersaurus were the same.[24]
Nomina dubia (doubtful species)
* D. lacustris is a nomen dubium, named by Marsh in 1884, from remains of a smaller animal from Morrison, Colorado.[25] These remains are now believed to have been from an immature animal, rather than from a separate species.[26]
Paleobiology
Due to a wealth of skeletal remains, Diplodocus is one of the best-studied dinosaurs. Many aspects of its lifestyle have been subjects of various theories over the years.
Habitat
Marsh and then Hatcher[27] assumed the animal was aquatic, because of the position of its nasal openings at the apex of the cranium. Similar aquatic behavior was commonly depicted for other large sauropods such as Brachiosaurus and Apatosaurus. However, a 1951 study by Kenneth A. Kermack indicates that sauropods probably could not have breathed through their nostrils when the rest of the body was submerged, as the water pressure on the chest wall would be too great.[28] Since the 1970s, general consensus has the sauropods as firmly terrestrial animals, browsing on trees. However, a more recent theory[clarification needed][citation needed] suggests a likely riparian habitat for Diplodocus.
Posture
The depiction of Diplodocus posture has changed considerably over the years. For instance, a classic 1910 reconstruction by Oliver P. Hay depicts two Diplodocus with splayed lizard-like limbs on the banks of a river. Hay argued that Diplodocus had a sprawling, lizard-like gait with widely-splayed legs,[30] and was supported by Gustav Tornier. However, this hypothesis was put to rest by W. J. Holland, who demonstrated that a sprawling Diplodocus would have needed a trench to pull its belly through.[31]
Later, diplodocids were often portrayed with their necks held high up in the air, allowing them to graze from tall trees. More recently, scientists have argued that the heart would have had trouble sustaining sufficient blood pressure to oxygenate the brain. Furthermore, more recent studies have shown that the structure of the neck vertebrae would not have permitted the neck to bend far upwards.[32][33]
As with the related genus Barosaurus, the very long neck of Diplodocus is the source of much controversy among scientists. A 1992 Columbia University study of Diplodocid neck structure indicated that the longest necks would have required a 1.6 ton heart — a tenth of the animal's body weight. The study proposed that animals like these would have had rudimentary auxiliary 'hearts' in their necks, whose only purpose was to pump blood up to the next 'heart'.[4]
While the long neck has traditionally been interpreted as a feeding adaptation, a recent study[34] suggests that the oversized neck of Diplodocus and its relatives may have been primarily a sexual display, with any other feeding benefits coming second.
Diet
Diplodocus has highly unusual teeth compared to other sauropods. The crowns are long and slender, elliptical in cross-section, while the apex forms a blunt triangular point.[8] The most prominent wear facet is on the apex, though unlike all other wear patterns observed within sauropods, Diplodocus wear patterns are on the labial (cheek) side of both the upper and lower teeth.[8] What this means is Diplodocus and other diplodocids had a radically different feeding mechanism than other sauropods. Unilateral branch-stripping is the most likely feeding behavior of Diplodocus,[35][36][37] as it explains the unusual wear patterns of the teeth (coming from tooth-food contact). In unilateral branch stripping, one tooth row would have been used to strip foliage from the stem, while the other would act as a guide and stabilizer. With the elongated preorbital (in-front of the eyes) region of the skull, longer portions of stems could be stripped in a single action.[8] Also the palinal (backwards) motion of the lower jaws could have contributed two significant roles to feeding behaviour: 1) an increased gape, and 2) allowed fine adjustments of the relative positions of the tooth rows, creating a smooth stripping action.[8]
With a laterally and dorsoventrally flexible neck, and the possibility of using its tail and rearing up on its hind limbs (tripodal ability), Diplodocus would have had the ability to browse at many levels (low, medium, and high), up to approximately 10 metres (33 ft) from the ground.[38] The neck's range of movement would have also allowed the head to graze below the level of the body, leading some scientists to speculate on whether Diplodocus grazed on submerged water plants, from riverbanks. This concept of the feeding posture is supported by the relative lengths of front and hind limbs. Furthermore, its peglike teeth may have been used for eating soft water plants.[32]
Other anatomical aspects
The head of Diplodocus has been widely depicted with the nostrils on top due to the position of the nasal openings at the apex of the skull. There has been speculation over whether such a configuration meant that Diplodocus may have had a trunk.[39] A recent study[40] surmised there was no paleoneuroanatomical evidence for a trunk. It noted that the facial nerve in an animal with a trunk, such as an elephant, is large as it innervates the trunk. The evidence suggests that the facial nerve is very small in Diplodocus. Studies by Lawrence Witmer (2001) indicated that, while the nasal openings were high on the head, the actual, fleshy nostrils were situated much lower down on the snout.[41]
Recent discoveries have suggested that Diplodocus and other diplodocids may have had narrow, pointed keratinous spines lining their back, much like those on an iguana.[42][43] This radically different look has been incorporated into recent reconstructions, notably Walking with Dinosaurs. It is unknown exactly how many diplodocids had this trait, and whether it was present in other sauropods.
Reproduction and growth
While there is no evidence for Diplodocus nesting habits, other sauropods such as the titanosaurian Saltasaurus have been associated with nesting sites.[44][45] The titanosaurian nesting sites indicate that may have laid their eggs communally over a large area in many shallow pits, each covered with vegetation. It is possible that Diplodocus may have done the same. The documentary Walking with Dinosaurs portrayed a mother Diplodocus using an ovipositor to lay eggs, but it was pure speculation on the part of the documentary.
Following a number of bone histology studies, Diplodocus, along with other sauropods, grew at a very fast rate, reaching sexual maturity at just over a decade, though continuing to grow throughout their lives.[46][47][48] Previous thinking held that sauropods would keep growing slowly throughout their lifetime, taking decades to reach maturity.
Classification
Diplodocus is both the type genus of, and gives its name to Diplodocidae, the family to which it belongs.[25] Members of this family, while still massive, are of a markedly more slender build when compared with other sauropods, such as the titanosaurs and brachiosaurs. All are characterised by long necks and tails and a horizontal posture, with forelimbs shorter than hindlimbs. Diplodocids flourished in the Late Jurassic of North America and possibly Africa[14] and appear to have been replaced ecologically by titanosaurs during the Cretaceous.
A subfamily, Diplodocinae, was erected to include Diplodocus and its closest relatives, including Barosaurus. More distantly related is the contemporaneous Apatosaurus, which is still considered a diplodocid although not a diplodocine, as it is a member of the subfamily Apatosaurinae.[49][50] The Portuguese Dinheirosaurus and the African Tornieria have also been identified as close relatives of Diplodocus by some authors.[51][52]
The Diplodocoidea comprises the diplodocids, as well as dicraeosaurids, rebbachisaurids, Suuwassea,[49][50] Amphicoelias[52] and possibly Haplocanthosaurus,[53] and/or the nemegtosaurids.[7] This clade is the sister group to, Camarasaurus, brachiosaurids and titanosaurians; the Macronaria.[7][53] Together they comprise Neosauropoda; the largest, most diverse and successful group of sauropodomorph dinosaurs.
In popular culture
Diplodocus has been a famous and much-depicted dinosaur. Much of this has probably been due to its wealth of skeletal remains and former status as the longest dinosaur. However, the donation of many mounted skeletal casts by industrialist Andrew Carnegie to potentates around the world at the beginning of the twentieth century[54] did much to familiarize it to people worldwide. Casts of Diplodocus skeletons are still displayed in many museums worldwide, including an unusual D. hayi in the Houston Museum of Natural Science, and D. carnegiei in the Natural History Museum in London, the Natural Science Museum in Madrid, Spain, the Museum für Naturkunde in Berlin, Germany, the Senckenberg Museum in Frankfurt, Germany, the Field Museum of Natural History in Chicago and, of course, the original is still on display at the Carnegie Museum of Natural History in Pittsburgh. A mounted skeleton of D. longus is at the Smithsonian National Museum of Natural History in Washington, D.C., while a mounted skeleton of D. hallorum (formerly Seismosaurus), which may be the same as D. longus, can be found at the New Mexico Museum of Natural History and Science.
Diplodocus has been a frequent subject in dinosaur films, both factual and fictional. It was featured in the second episode of the award-winning BBC television series Walking with Dinosaurs. The episode "Time of the Titans" follows the life of a simulated Diplodocus 152 million years ago. The animated feature Fantasia features many sauropods in the Rite of Spring sequence, some narrower-headed ones of which may be Diplodocus. In literature, James A. Michener's book Centennial has a chapter devoted to Diplodocus, narrating the life and death of one individual.
Diplodocus is a commonly-seen figure in dinosaur toy and scale model lines. It has had two separate figures in the Carnegie Collection.
References
1. ^ a b c Simpson, John; Edmund Weiner (eds.) (1989). The Oxford English Dictionary (2nd edition ed.). Oxford: Oxford University Press. ISBN 0-19-861186-2.
2. ^ a b Pickett, Joseph P. et al. (eds.) (2000). The American Heritage Dictionary of the English Language (4th edition ed.). Boston: Houghton Mifflin Company. ISBN 0-395-82517-2.
3. ^ Christine C.E. & Peterson, F. (2004). "Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem—a synthesis". Sedimentary Geology 167, 309–355
4. ^ a b c Lambert D. (1993)The Ultimate Dinosaur Book ISBN 0-86438-417-3
5. ^ Wedel, M.J. and Cifelli, R.L. Sauroposeidon: Oklahoma's Native Giant. 2005. Oklahoma Geology Notes 65:2.
6. ^ a b Upchurch P, Barrett PM, Dodson P (2004). "Sauropoda". in Weishampel DB, Dodson P, Osmólska H. The Dinosauria (2nd Edition), University of California Press. pp. 305. ISBN 0-520-24209-2.
7. ^ a b c Upchurch P, Barrett PM, Dodson P (2004). "Sauropoda". in Weishampel DB, Dodson P, Osmólska H. The Dinosauria (2nd Edition), University of California Press. pp. 316. ISBN 0-520-24209-2.
8. ^ a b c d e Upchurch, P. & Barrett, P.M. (2000). Chapter 4: The evolution of sauropod feeding mechanism. IN: Evolution of Herbivory in Terrestrial Vertebrates ISBN 0-521-59449-9
9. ^ Stevens, K.A. & Parrish, M. (1999). "Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs". Science 284, 798–800
10. ^ Dodson, P., Behrensmeyer, A.K., Bakker, R.T., and McIntosh, J.S. (1980). Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation. Paleobiology 6:208–232.
11. ^ Paul, G.S. (1994). Big sauropods - really, really big sauropods. The Dinosaur Report, The Dinosaur Society Fall:12–13.
12. ^ Foster, J.R. (2003). Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. New Mexico Museum of Natural History and Science:Albuquerque, New Mexico. Bulletin 23.
13. ^ Coe, M.J., Dilcher, D.L., Farlow, J.O., Jarzen, D.M., and Russell, D.A. (1987). Dinosaurs and land plants. In: Friis, E.M., Chaloner, W.G., and Crane, P.R. (eds.). The Origins of Angiosperms and Their Biological Consequences. Cambridge University Press:New York, 225–258. ISBN 0521323576.
14. ^ a b Wilson JA (2005). "Overview of Sauropod Phylogeny and Evolution". in Rogers KA & Wilson JA(eds). The Sauropods:Evolution and Paleobiology, Indiana University Press. p. 15–49. ISBN 0-520-24623-3.
15. ^ Holland WJ (1915). "Heads and Tails: a few notes relating to the structure of sauropod dinosaurs.". Annals of the Carnegie Museum 9: 273–278.
16. ^ Myhrvold NP and Currie PJ (1997). "Supersonic sauropods? Tail dynamics in the diplodocids". Paleobiology 23: 393–409.
17. ^ Bonnan, M. F. (2003). "The evolution of manus shape in sauropod dinosaurs: implications for functional morphology, forelimb orientation, and phylogeny." Journal of Vertebrate Paleontology, 23: 595-613.
18. ^ Marsh OC. Principal characters of American Jurassic dinosaurs. Part I. American Journal of Science 3; 411–416 (1878).
19. ^ McIntosh (2005). "The Genus Barosaurus (Marsh)". in Carpenter, Kenneth and Tidswell, Virginia (ed.). Thunder Lizards: The Sauropodomorph Dinosaurs, Indiana University Press. pp. 38–77. ISBN 0-253-34542-1.
20. ^ Holland WJ. The skull of Diplodocus. Memoirs of the Carnegie Museum IX; 379–403 (1924).
21. ^ Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus." In Foster, J.R. and Lucas, S.G., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131-138.[1]
22. ^ Lucas S, Herne M, Heckert A, Hunt A, and Sullivan R. Reappraisal of Seismosaurus, A Late Jurassic Sauropod Dinosaur from New Mexico. The Geological Society of America, 2004 Denver Annual Meeting (November 7–10, 2004). Retrieved on 2007-05-24.
23. ^ Lucas, S.G., Spielman, J.A., Rinehart, L.A., Heckert, A.B., Herne, M.C., Hunt, A.P., Foster, J.R., and Sullivan, R.M. (2006). "Taxonomic status of Seismosaurus hallorum, a Late Jurassic sauropod dinosaur from New Mexico". in Foster, J.R., and Lucas, S.G.. Paleontology and Geology of the Upper Morrison Formation, New Mexico Museum of Natural History and Science (bulletin 36). pp. 149–161. ISSN 1524-4156.
24. ^ Lovelace, David M.; Hartman, Scott A.; and Wahl, William R. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional 65 (4): 527–544.
25. ^ a b Marsh, O.C. 1884. Principal characters of American Jurassic dinosaurs. Part VII. On the Diplodocidae, a new family of the Sauropoda. American Journal of Science 3: 160–168.
26. ^ Upchurch, P., Barrett, P.M., and Dodson, P. (2004). "Sauropoda." In D. B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria (2nd edition). University of California Press, Berkeley 259–322.
27. ^ Hatcher JB. "Diplodocus (Marsh): Its osteology, taxonomy, and probable habits, with a restoration of the skeleton,". Memoirs of the Carnegie Museum, vol. 1 (1901), pp. 1–63
28. ^ Kermack, Kenneth A. (1951). "A note on the habits of sauropods". Annals and Magazine of Natural History 12 (4): 830–832.
29. ^ Hay, O. P., 1910, Proceedings of the Washington Academy of Sciences, vol. 12,, pp. 1–25
30. ^ Hay, Dr. Oliver P., "On the Habits and Pose of the Sauropod Dinosaurs, especially of Diplodocus." The American Naturalist, Vol. XLII, Oct. 1908
31. ^ Holland, Dr. W. J., "A Review of Some Recent Criticisms of the Restorations of Sauropod Dinosaurs Existing in the Museums of the United States, with Special Reference to that of Diplodocus carnegiei in the Carnegie Museum", The American Naturalist, 44:259–283. 1910.
32. ^ a b Stevens KA, Parrish JM (2005). "Neck Posture, Dentition and Feeding Strategies in Jurassic Sauropod Dinosaurs". in Carpenter, Kenneth and Tidswell, Virginia (ed.). Thunder Lizards: The Sauropodomorph Dinosaurs, Indiana University Press. pp. 212–232. ISBN 0-253-34542-1.
33. ^ Upchurch, P, et al. (2000). ""Neck Posture of Sauropod Dinosaurs"" (PDF). Science 287, 547b (2000); DOI: 10.1126/science.287.5453.547b. Retrieved on 2006-11-28.
34. ^ Senter, P. "Necks for Sex: Sexual Selection as an Explanation for Sauropod Neck Elongation". Journal of Zoology, 2006
35. ^ Norman, D.B. (1985). "The illustrated Encyclopedia of Dinosaurs". London: Salamander Books Ltd
36. ^ Dodson, P. (1990). Sauropod paleoecology. IN: "The Dinosauria" 1st Edition, (Eds. Weishampel, D.B., Dodson, P. & Osmólska, H.)
37. ^ Barrett, P.M. & Upchurch, P. (1994). Feeding mechanisms of Diplodocus. Gaia 10, 195–204
38. ^ Barrett, P.M. & Upchurch, P. (2005). Sauropodomorph Diversity through Time, Paleoecological and Macroevolutionary Implications. IN: "The Sauropods: Evolution and Paleobiology" (Eds. Curry, K. C.)
39. ^ Bakker, Robert T. (1986) The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. New York: Morrow.
40. ^ Knoll, F., Galton, P.M., López-Antoñanzas, R. (2006). "Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus." Geobios, 39: 215–221
41. ^ Lawrence M. Witmer et al., "Nostril Position in Dinosaurs and other Vertebrates and its Significance for Nasal Function." Science 293, 850 (2001)
42. ^ Czerkas, S. A. (1993). "Discovery of dermal spines reveals a new look for sauropod dinosaurs." Geology 20, 1068–1070
43. ^ Czerkas, S. A. (1994). "The history and interpretation of sauropod skin impressions." In Aspects of Sauropod Paleobiology (M. G. Lockley, V. F. dos Santos, C. A. Meyer, and A. P. Hunt, Eds.), Gaia No. 10. (Lisbon, Portugal).
44. ^ Walking on Eggs: The Astonishing Discovery of Thousands of Dinosaur Eggs in the Badlands of Patagonia, by Luis Chiappe and Lowell Dingus. 19 June 2001, Scribner
45. ^ Grellet-Tinner, Chiappe, & Coria, "Eggs of titanosaurid sauropods from the Upper Cretaceous of Auca Mahuevo (Argentina)", Can. J. Earth Sci. 41(8): 949–960 (2004)
46. ^ Sander, P. M. (2000). "Long bone histology of the Tendaguru sauropods: Implications for growth and biology". Paleobiology 26, 466–488
47. ^ Sander, P. M., N. Klein, E. Buffetaut, G. Cuny, V. Suteethorn, and J. Le Loeuff (2004). "Adaptive radiation in sauropod dinosaurs: Bone histology indicates rapid evolution of giant body size through acceleration". Organisms, Diversity & Evolution 4, 165–173
48. ^ Sander, P. M., and N. Klein (2005). "Developmental plasticity in the life history of a prosauropod dinosaur". Science 310 1800–1802
49. ^ a b Taylor, M.P. & Naish, D. 2005. The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda). PaleoBios 25(2): 1–7. (download here)
50. ^ a b Harris, J.D. 2006. The significance of Suuwassea emiliae (Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution. Journal of Systematic Palaeontology 4(2): 185–198.
51. ^ Bonaparte, J.F. & Mateus, O. 1999. A new diplodocid, Dinheirosaurus lourinhanensis gen. et sp. nov., from the Late Jurassic beds of Portugal. Revista del Museo Argentino de Ciencias Naturales. 5(2):13–29. (download here)
52. ^ a b Rauhut, O.W.M., Remes, K., Fechner, R., Cladera, G., & Puerta, P. 2005. Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature 435: 670–672.
53. ^ a b Wilson, J. A., 2002, "Sauropod dinosaur phylogeny: critique and cladistica analysis". Zoological Journal of the Linnean Society 136: 217–276.
54. ^ Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking The Mystery of the Dinosaurs and Their Extinction. New York: William Morrow. pp. 203. ISBN 0140100555.
External links
- Diplodocus in the Dino Directory
- Museo Nacional de Ciencias Naturales, Madrid
- Diplodocus Marsh, by J.B. Hatcher 1901 - Its Osteology, Taxonomy, and Probable Habits, with a Restoration of the Skeleton. Memoirs of the Carnegie Museum, Volume 1, Number 1, 1901. Full text, Free to read.
- Carnegie Museum of Natural History - History
- Skeletal restorations of diplodocids including D. carnegii, D. longus, and D. hallorum, from Scott Hartman's Skeletal Drawing website.
- Chapter 5: The Amphibious Dinosaurs - Brontosaurus, Diplodocusw, Etc. Sub-Order Opisthocœlia (Cetiosauria or Sauropoda by W. D. Matthew, who is credited amongst other accomplishments as authorship of the family Dromaeosauridae, and former Curator of Vertebrate Paleontology at the American Museum of Natural History in New York; Originally published in 1915
Retrieved from "http://en.wikipedia.org/"
All text is available under the terms of the GNU Free Documentation License

{kind=link}