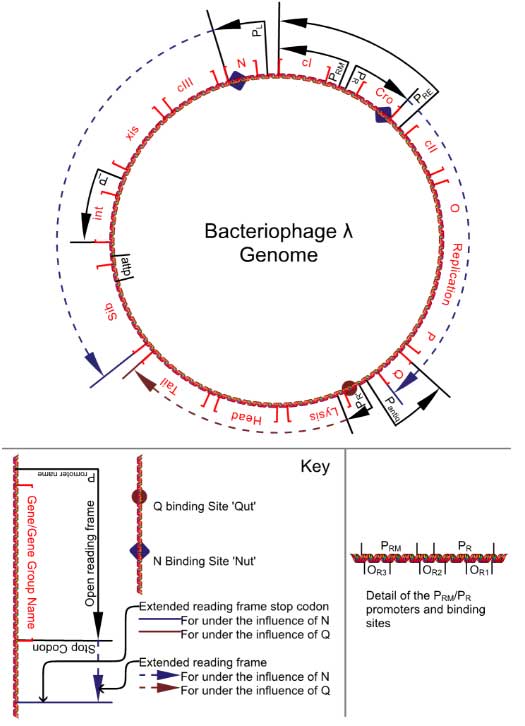
Enterobacteria phage λ (lambda phage, coliphage λ) is a temperate bacteriophage that infects Escherichia coli. Lambda phage is a virus particle consisting of a head, containing double-stranded linear DNA as its genetic material, and a tail that can have tail fibers. The phage particle injects its DNA into its host through the tail, and the phage will then usually enter the lytic pathway where it replicates its DNA, degrades the host DNA and hijacks the cell's replication, transcription and translation mechanisms to produce as many phage particles as cell resources allow. When cell resources are depleted, the phage will lyse (break open) the host cell, releasing the new phage particles. However, under certain conditions, the phage DNA may integrate itself into the host cell chromosome in the lysogenic pathway. In this state, the λ DNA is called a prophage and stays resident within the host's genome without apparent harm to the host, which can be termed a lysogen when a prophage is present. The prophage is duplicated with every subsequent cell division of the host. The phage genes expressed in this dormant state code for proteins that repress expression of other phage genes. These proteins are broken down when the host cell is under stress, resulting in the expression of the repressed phage genes. Stress can be from starvation, poisons (like antibiotics), or other factors that can damage or destroy the host. In response to stress, the activated prophage is excised from the DNA of the host cell by one of the newly expressed gene products and enters its lytic pathway. Lambda phage was discovered by Esther Lederberg in 1950.[1] It has been used heavily as a model organism, and has been a rich source for useful tools in molecular biology. Uses include its application as a vector for the cloning of recombinant DNA, the use of its site-specific recombinase, int, for the shuffling of cloned DNAs by the 'Gateway' method, and the application of its Red operon, including the proteins Red alpha (also called 'exo'), beta and gamma in the DNA engineering method called recombineering. In the following page, genes will be written in italics and their associated proteins in Roman. For instance, cI refers to the gene, while cI is the resulting protein encoded by that gene. Anatomy The virus particle consists of a head and a tail that can have tail fibres. The head contains 48,490 base pairs of double-stranded, linear DNA flanked by 12-base-pair, single-stranded segments that make up the two strands of the cos site. In its circular form in the host cytoplasm, the phage genome therefore is 48,502 base pairs in length. The prophage exists as a linear section of DNA inserted into the host chromosome. Lifecycle Infection 1. Bacteriophage Lambda binds to the target E. coli cell, the J protein in the tail tip interacting with the lamB gene product of E. coli, a porin molecule which is part of the maltose operon.
This occurs without the N protein interacting with the DNA; the protein instead binds to the freshly transcribed mRNA. Nut sites contain 3 conserved "boxes," of which only BoxB is essential. 1. The boxB RNA sequences are located close to the 5' end of the pL and pR transcripts. When transcribed, each sequence forms a hairpin loop structure that the N protein can bind to.
This is the lifecycle that the phage follows following most infections, where the cII protein does not reach a high enough concentration due to degradation, so does not activate its promoters. 1. The 'late early' transcripts continue being written, including xis, int, Q and genes for replication of the lambda genome (OP). Cro dominates the repressor site (see "Repressor" section), repressing synthesis from the PRM promoter.
Rightward transcription expresses the O, P and Q genes. O and P are responsible for initiating replication, and Q is another antiterminator which allows the expression of head, tail and lysis genes from PR’. Lytic replication 1. For the first few replication cycles, the lambda genome undergoes θ replication (circle-to-circle).
Q is similar to N in its effect: Q binds to RNA polymerase in Qut sites and the resulting complex can ignore terminators, however the mechanism is very different; the Q protein first associates with a DNA sequence rather than an mRNA sequence[3]. 1. The Qut site is very close to the PR’ promoter, close enough that the σ factor has not been released from the RNA polymerase holoenzyme. Part of the Qut site resembles the -10 Pribnow box, causing the holoenzyme to pause.
Leftward transcription expresses the gam, red, xis and int genes. Gam and red proteins are involved in recombination. Gam is also important in that it inhibits the host RecBCD nuclease from degrading the 3’ ends in rolling circle replication. Int and xis are integration and excision proteins which are vital to lysogeny. xis and int regulation of insertion and excision 1. xis and int are found on the same piece of mRNA, so approximately equal concentrations of xis and int proteins are produced. This results (initially) in the excision of any inserted genomes from the host genome.
This is the lifecycle that the phage follows after a small number of infections in specific conditions, where the cII protein reaches a high enough concentration due to stabilisation and lack of degradation, and so activates its promoters. 1. The 'late early' transcripts continue being written, including xis, int, Q and genes for replication of the lambda genome.
The integration of phage λ takes place at a special attachment site in the bacterial and phage genomes, called attλ. The sequence of the bacterial att site is called attB, between the gal and bio operons, and consists of the parts B-O-B', whereas the complementary sequence in the circular phage genome is called attP and consists of the parts P-O-P'. The integration itself is a sequential exchange (see genetic recombination) via a Holliday junction and requires both the phage protein Int and the bacterial protein IHF (integration host factor). Both Int and IHF bind to attP and form an intasome, a DNA-protein-complex designed for site-specific recombination of the phage and host DNA. The original B-O-B' sequence is changed by the integration to B-O-P'-phage DNA-P-O-B'. The phage DNA is now part of the host's genome. Maintenance of lysogeny * Lysogeny is maintained solely by cI. cI represses transcription from PL and PR while upregulating and controlling its own expression from PRM. It is therefore the only protein expressed by lysogenic phage.
The classic induction of a lysogen involved irradiating the infected cells with UV light. Any situation where a lysogen undergoes DNA damage or the SOS response of the host is otherwise stimulated leads to induction. 1. The host cell, containing a dormant phage genome, experiences DNA damage due to a high stress environment, and starts to undergo the SOS response.
Schematic representation of the insertion of the bacteriophage lambda. Note how sib is displaced by the recombination event from the N extended PL promoter open reading frame. (*) 1. The phage genome is still inserted in the host genome and needs excision for DNA replication to occur. The sib section beyond the normal PL promoter transcript is, however, no longer included in this reading frame (see diagram).
Protein function overview cro; (Control of Repressor's Operator) Transcription inhibitor, binds OR3, OR2 and OR1 (affinity OR3 > OR2 = OR1, i.e. preferentially binds OR3). At low concentrations blocks the pRM promoter (preventing cI production). At high concentrations downregulates its own production through OR2 and OR1 binding. No cooperative binding (c.f. below for cI binding) cI; (Clear 1) Transcription inhibitor, binds OR1, OR2 and OR3 (affinity OR1 > OR2 = OR3, i.e. preferentially binds OR1). At low concentrations blocks the pR promoter (preventing cro production). At high concentrations downregulates its own production through OR3 binding. Binding of cI at OR1 stimulates an almost simultaneous cI binding to OR2 via cooperative binding (via cI C terminal domain interactions) N terminal domain of cI on OR2 tightens the binding of RNA polymerase to pRM and hence stimulate its own transcription. Repressor also inhibits transcription from the pL promoter. Susceptible to cleavage by RecA* in cells undergoing the SOS response. cII; (Clear 2) Transcription activator. Activates transcription from the pAQ, pRE and pI promoters. Low stability due to susceptibility to cellular HflB (FtsH) proteases (especially in healthy cells and cells undergoing the SOS response). cIII;(Clear 3) HflB (FtsH) binding protein, protects cII from degradation by proteases. N; (aNtiterminator) RNA binding protein and RNA polymerase cofactor, binds RNA (at Nut sites) and transfers onto the nascent RNApol that just transcribed the nut site. This RNApol modification prevents its recognition of termination sites, so normal RNA polymerase termination signals are ignored and RNA synthesis continues into distal phage genes. Q; DNA binding protein and RNApol cofactor, binds DNA (at Qut sites) and transfers onto the initiating RNApol. This RNApol modification alters its recognition of termination sequences, so normal ones are ignored; special Q termination sequences some 20,000 bp away are effective. xis; (eXcISion) excisionase and Int protein regulator, manages excision and insertion of phage genome into the host's genome. int; (INTegration) Int protein, manages insertion of phage genome into the host's genome. In Conditions of low int concentration there is no effect. If xis is low in concentration and int high then this leads to the insertion of the phage genome. If xis and int have high (and approximately equal) concentrations this leads to the excision of phage genomes from the host's genome. A, B, C, D, E, F, Z, U, V, G, T, H, M, L, K, I, J [Shown on diagram as head and tail, A-F code for phage head genes, Z-J code for phage tail genes. The order shown here is as found on the genome, reading in a clockwise direction]; structural proteins, self assemble with the phage genome into daughter phage particles. S, R [Shown on diagram as lysis. The order shown here is as found on the genome, reading in a clockwise direction]; cause the host cell to undergo lysis at high enough concentrations. OP [Shown on diagram as O replication P]; DNA replication functions, promotes the specific replication of only the phage genome. sib [not a protein, but a vital conserved DNA sequence]; Forms a stable hairpin loop structure in transcribed mRNA beyond int. Attracts degradation of mRNA by RNAaseIII. attP [not a protein, but a conserved DNA sequence]; point of action of Int and Xis in integration and excision of the phage genome into the host's genome. Corresponding attB found in the host's genome at the point of insertion. Repressor The repressor found in the phage lambda is a notable example of the level of control possible over gene expression by a very simple system. It forms a 'binary switch' with two genes under mutually exclusive expression, as discovered by Barbara J. Meyer.[4] The lambda repressor gene system consists of (from left to right on the chromosome): * cI gene The lambda repressor is a dimer also known as the cI protein. It binds DNA in the helix-turn-helix binding motif. It regulates the transcription of the cI protein and the Cro protein. The life cycle of lambda phages is controlled by cI and Cro proteins. The lambda phage will remain in the lysogenic state if cI proteins predominate, but will be transformed into the lytic cycle if cro proteins predominate. The cI dimer may bind to any of three operators, OR1, OR2, and OR3, in the order OR1 = OR2 > OR3. Binding of a cI dimer to OR1 enhances binding of a second cI dimer to OR2, an effect called cooperativity. Thus, OR1 and OR2 are almost always simultaneously occupied by cI. However, this does not increase the affinity between cI and OR3, which will be occupied only when the cI concentration is high. This cooperative action is shown by the relative affinity of the repressor for the native sequences individually, which is OR1 > OR2 = OR3; different from the actual order of binding. * In the absence of cI proteins, the cro gene may be transcribed.
The gene regulatory circuitry of phage λ is among the best-understood circuits at the mechanistic level. This circuitry involves several interesting regulatory behaviors. An infected cell undergoes a decision between two alternative pathways, the lytic and lysogenic pathways. If the latter is followed, the lysogenic state is established and maintained. While this state is highly stable, it can switch to the lytic pathway in the process of prophage induction, which occurs when the host SOS response is triggered by DNA damage.[5] An important distinction here is that between the two decisions; lysogeny and lysis on infection, and continuing lysogeny or lysis from a prophage. The latter is determined solely by the activation of RecA in the SOS response of the cell, as detailed in the section on induction. The former will also be affected by this; a cell undergoing an SOS response will always by lysed, as no cI protein will be allowed to build up. However, the initial lytic/lysogenic decision on infection is also dependent on the cII and cIII proteins. Simplistically, in cells with sufficient nutrients, protease activity is high, which breaks down cII.[6] This leads to the lytic lifestyle. In cells with limited nutrients, protease activity is low, making cII stable. This leads to the lysogenic lifestyle. cIII appears to stabilise cII, both directly and by acting as a competitive inhibitor to the relevant proteases. This means that a cell "in trouble", i.e. lacking in nutrients and in a more dormant state, is more likely to lysogenise. This would be selected for because the phage can now lie dormant in the bacterium until it falls on better times, and so the phage can create more copies of itself with the additional resources available and with the more likely proximity of further infectable cells. Realistically, a full biophysical model for lambda's lysis-lysogeny decision remains to be developed. Computer modeling and simulation suggest that random processes during infection drive the selection of lysis or lysogeny within individual cells.[7] However, recent experiments suggest that physical differences among cells, that exist prior to infection, predetermine whether a cell will lyse or become a lysogen.[8] See also * molecular weight size marker
1. ^ Esther Lederberg, "Lysogenicity in Eescherichia coli strain K-12, Microbial Genetics Bulletin, v.1, pp. 5-8 (January 1950); followed by "Lysogenicity in E. coli K-12", Genetics, v.36, p. 560 (1951) (abstract).
* Esther Lederberg, "Lysogenicity in Escherichia coli strain K-12", Microbial Genetics Bulletin, v.1, pp. 5–8 (January 1950); followed by "Lysogenicity in E. coli K-12", Genetics, v.36, p. 560 (1951) (abstract) (discovery reported). Retrieved from "http://en.wikipedia.org/"
|

{kind=link}