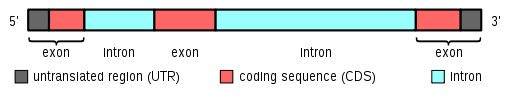
An exon is a nucleic acid sequence that is represented in the mature form of an RNA molecule after either portions of a precursor RNA (introns) have been removed by cis-splicing or when two or more precursor RNA molecules have been ligated by trans-splicing. The mature RNA molecule can be a messenger RNA or a functional form of a non-coding RNA such as rRNA or tRNA. Depending on the context, exon can refer to the sequence in the DNA or its RNA transcript. History The term exon was coined by American biochemist Walter Gilbert in 1978: "The notion of the cistron… must be replaced by that of a transcription unit containing regions which will be lost from the mature messenger – which I suggest we call introns (for intragenic regions) – alternating with regions which will be expressed – exons."[1] This definition was originally made for protein-coding transcripts that are spliced before being translated. The term later came to include sequences removed from rRNA[2] and tRNA,[3] and it also was used later for RNA molecules originating from different parts of the genome that are then ligated by trans-splicing.[4] Function In many genes, each exon contains part of the open reading frame (ORF) that codes for a specific portion of the complete protein. However, the term exon is often misused to refer only to coding sequences for the final protein. This is incorrect, since many noncoding exons are known in human genes (Zhang 1998).
Some of the exons will be wholly or part of the 5' untranslated region (5' UTR) or the 3' untranslated region (3' UTR) of each transcript. The untranslated regions are important for efficient translation of the transcript and for controlling the rate of translation and half life of the transcript. Furthermore, transcripts made from the same gene may not have the same exon structure since parts of the mRNA could be removed by the process of alternative splicing. Some mRNA transcripts have exons with no ORF's and thus are sometimes referred to as non-coding RNA. Exonization is the creation of a new exon, as result of mutations in intronic sequences.[5] Polycistronic messages have multiple ORF's in one transcript and also have small regions of untranslated sequence between each ORF. Experimental approaches that utilize exons Exon trapping or 'gene trapping' is a molecular biology technique that exploits the existence of the intron-exon splicing to find new genes. The first exon of a 'trapped' gene splices into the exon that is contained in the insertional DNA. This new exon contains the ORF for a reporter gene that can now be expressed using the enhancers that control the target gene. A scientist knows that a new gene has been trapped when the reporter gene is expressed. Splicing can be experimentally modified so that targeted exons are excluded from mature mRNA transcripts by blocking the access of splice-directing small nuclear ribonucleoprotein particles (snRNPs) to pre-mRNA using Morpholino antisense oligos.[6] This has become a standard technique in developmental biology. Morpholino oligos can also be targeted to prevent molecules that regulate splicing (e.g. splice enhancers, splice suppressors) from binding to pre-mRNA, altering patterns of splicing. See also * Eukaryotic gene example
1. ^ Gilbert W (February 1978). "Why genes in pieces?". Nature 271 (5645): 501. doi:10.1038/271501a0. PMID 622185.
* Gilbert W (February 1978). "Why genes in pieces?". Nature 271 (5645): 501. doi:10.1038/271501a0. PMID 622185.
* Exon-intron graphic maker Retrieved from "http://en.wikipedia.org/"
|

{kind=link}