|
|
Elasmotherium sibiricum, Photo: Michael Lahanas Cladus: Eukaryota Name Elasmotherium sibiricum J. Fischer, 1809 Vernacular Names Elasmotherium ("Thin Plate Beast"), more informally sometimes called the Steppe Rhiniceros, is an extinct genus of giant rhinoceros endemic to Eurasia during the Late Pliocene through the Pleistocene, documented from 2.6 mya to as late as 50,000 years ago, possibly later, in the Late Pleistocene, an approximate span of slightly less than 2.6 million years. Three species are recognised. The best known, E. sibiricum was the size of a mammoth and is thought to have borne a large, thick horn on its forehead, which it undoubtedly used for the same purposes as all the other horned savanna giants then and now: defense, attracting mates, driving away competitors, sweeping snow from the grass in winter and digging for water and plant roots. Like all rhinoceroses, elasmotheres were herbivorous. Unlike any others, its high-crowned molars were ever-growing. Its legs were longer than those of other rhinos and were designed for galloping, giving it a horse-like gait. The Russian paleontologists of the 19th century who discovered and named the initial fossils were influenced by ancient legends of a huge unicorn roaming the steppes of Siberia. To date no evidence either contradicts or confirms the possibility that E. survived into legendary times.
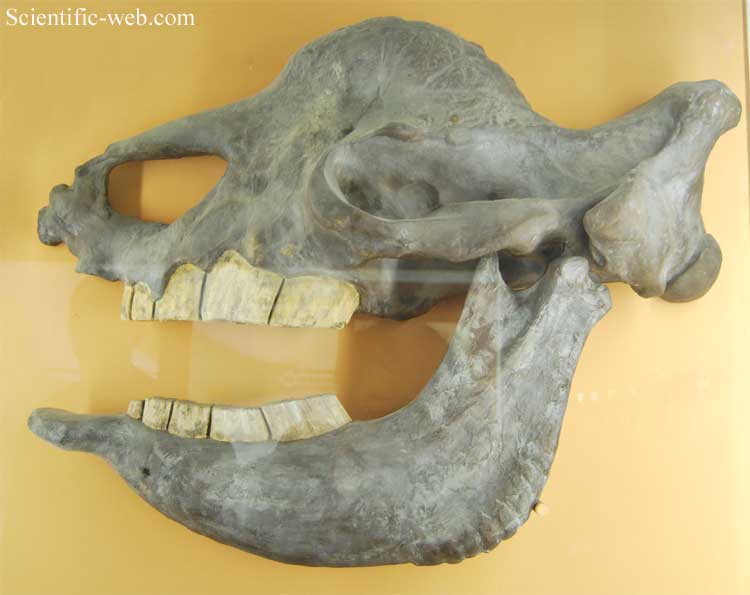
The fossil received its name from Johann Fischer von Waldheim,[2] the Dirécteur Perpétuel of the Natural History Museum, Moscow University, at a presentation before the Societé Impériale des Naturalistes in 1808. The next year in the Mémoires of the society he reported what he had said in the Programme d'invitation:[3] "L'elasmotherium est un animal à tête elongée sans dents incisives et sans canines, de chaque coté 5 molaires à lames contournées."[4] Then he noted on his derivation of the name: "Du mot grec ἐλασμος, lamelle, pour designer la forme lamelleuse des dents molaires."[5] All he had before him was one lower jaw donated to the museum by Yekaterina Romanovna Vorontsova-Dashkova, which he named Elasmotherium sibiricum, lamenting that it was the sole species of which he knew. The molars, the only teeth in the jaw, had formed in layers like tree rings, except the "rings," or lamellae, were highly corrugated. The edges in the grinding surface were elaborately sinuous to better break down the grasses on which the animal fed. Description
Morphology The most reconstructed species is perhaps E. sibiricum by generations of scientists working at the Paleontological Museum in Moscow and elsewhere in Russia. The majority of the fossils fall or have fallen within their national jurisdiction. The dimensions and morphology of the various reconstructions vary considerably. They are for the most part estimating the gross details from the minutiae. However, they all agree on the general order of magnitude, that sibiricum was comparable to a Mammoth and was rather larger than the contemporary Woolly Rhinoceros. E. sibiricum had a measured shoulder height of approximately 2 metres (6.6 ft). To it, however, must be added the height of a massive hump anchored on the fin-like transverse processes extending from the top of the cervical vertebrae, a maximum of 53 centimetres (1.74 ft). The total height then was in excess of 253 centimetres (8.30 ft). The measured length of sibiricum (from a nearly complete skeleton found at Gaevskaya) is 4.5 metres (15 ft). Extrapolation from the greater size of caucasicum molars obtains a length of 5 metres (16 ft)—5.2 metres (17 ft) for caucasicum.[6] The science of estimation is more advanced than it was when the 19th century estimates and reconstructions were made. A 1986 study of mammalian communities by Serge Legendre correlated body weight with the M1 area, which, in the code of dental formulae, means the area of the first lower molar. A correlation typically is expressed by a curve in a graph showing two (in this case) quantitative variables on a modified Cartesian coordinate system. The data is graphed as a scatter of points to which a smooth curve is then fitted in regression analysis. The curve fitted by Legendre for body weight of Perissodactyla in this case is the function ln x = 1.564ln y + 3.267 Skull with reconstructed horn where x is the weight, y is the M1 area (also called the ml area, the length of the molar grinding surface times its width), and the two numbers are constants applying only to perissodactyla.[7] According to Legendre's formula, sibiricum had a mass of over 4,000 kilograms (3.9 long tons; 4.4 short tons); caucasicum, 5,000 kilograms (4.9 long tons; 5.5 short tons).[8] These weights place Elasmotherium in the "really huge" category of all Rhinocerotidae and therefore the animal was "strongly brachyopodial;" that is, they required feet of large contact area to prevent sinking into the soil.[9] The feet were unguligrade, the front larger than the rear: purely tridactyl[10] on Digits II-IV in the rear, but with an extra vestigial digit, I, in the front.[6] Herbivores can be divided into two general groups on the basis of nutrition, which grade into each other morphologically: "foregut fermenters" and "hind-gut fermenters." The border region is correlated to bulk: up to 600 kilograms (1,300 lb) – 1,200 kilograms (2,600 lb) are the former; over it, the latter. In foregut fermentation the animal must "browse" to select the most nutritious plants and then ruminate to make up for the shorter digestive tract. The hind-gut fermenters are "bulk-feeders:" they ingest large quantities of low-nutrient food, which they process for a longer time in a much longer intestine. The main food in that category is grass, indicating that Elasmotherium, like the elephants, was probably a grassland "grazer" moving over long distances to take advantage of the growth phases of grass in different regions.[11] The standard is not without exception, as Indricotherium, the largest land mammal ever, with a weight of 15–20 tons, subsisted by browsing the treetops. There are also paleontological indications of the grazing. In general, the normal position of the head can be determined by the angle between a vertical plane coinciding with the occiput of the cranium, which is always vertical, and a plane through the base of the cranium. A right or acute angle would indicate a head held high for browsing leaves at various heights. Elasmotherium had the most obtuse angle of the Rhinocerotids. It could only reach the lowest levels and therefore must have grazed habitually.[12] This morphological feature favors the identification of the one-horned beast depicted in Rouffignac Cave (shown in this article) as Elasmotherium and lends some validity to the bison-like restorations based on it. Dentition A third type of evidence that Elasmotherium was a grazer is that of tooth wear and morphology. Like all Rhinocerotidae, E. had cheek teeth evolved for herbivory: two premolars and three molars (originally taken for five molars), no incisors, no canines.[13] Where some of the browsers kept the incisors in the form of tusks, E, had instead a spoon-like symphysis, or tip, of the lower mandible and a rostrum, or beak, of the upper, which served as a bony basis for a soft-tissue labial grasping and tearing mechanism.[14] Grass, a very tough, fibrous material, contains phytoliths, microscopic granules mainly of silica, which act as sandpaper on the molars of grazers. Their response in geologic time is to evolve cheek teeth with large crowns (hypsodonty). There appears to be a correlation between grazing and hypsodonty:[15] "As a general rule, extant herbivores with low-crowned teeth are predominantly browsers and species with high-crowned teeth are predominantly grazers." Vladimir Onufryevich Kovalevsky first proposed a connection between hypsodonty and grazing for horses in 1873.[16] Since then the concept has been expanded to all mammalian grazers at any time and has further been elaborated into hypsodonty or proto-hypsodonty and hypselodonty or euhypsodonty.[17] The euhypsodonts, of which, surprisingly for its bulk, E. was one,[18] have ever-growing high-crowned teeth. Most other examples are to be found among diminutive mammals such as Rodentia, which already casts doubt on the correlation, as they do not generally graze grass. Teeth form from the top down through the deposition of enamel on a cement core by formative soft tissue in the jaw. The enamel of hypsodont Perissodactyla is highly rugose rather than sharp. In brachydont species, such as humans, when the crown is complete, the roots are deposited and finally the completed tooth erupts. Hypselodonty is a condition of tooth eruption and continued crown formation before a delayed root formation. In its most developed variety, the roots never form. Only rare fossils of E. show any sign of a root, and that on a premolar. No molars have roots, or, in the terminology of some, the roots are "open." Habitats If all grazers are hypsodont, not all hypsodonts are grazers. The supposed correlation between grass-eating and hypsodonty proved difficult to support in a number of instances. Koenigswald, for example, pointed out that hypsodonty had occurred among the Gondwanatheria of the Mesozoic, a group of mammals so primitive that he describes their cheek teeth as "molariform," as they are neither clearly molars nor premolars.[19] Molariform hypsodonty "cannot be correlated with a grass diet, since grasses were not present." Instead he suggests for his example, Sudamerica ameghinioi, that it lived a "semiaquatic and perhaps a burrowing way of life." Modern species provide many examples from beavers to hippopotamuses. Attempts have been made to link the wear on E. teeth to grazing. In 1938, H.E. Wood, a Rhinocerotid tooth specialist, pointed out that interproximal wear, or loss of tooth surface between teeth, due to abrasion during mastication, of E. is similar to that of the White Rhinoceros, the only remaining Rhinocerotid grazer, which has hypsodont teeth.[20] Data such as this lead to an intuitive concept of some sort of correlation between grass-eating and hypsodonty, but it has been difficult to isolate mathematically. A 2007 study by Mendoza and Palmqvist compared the habitats and diets of 134 species of living ungulates, for which this data is known, both Artiodactyla and Perissodactyla, against "thirty-two craniodental measurements" to discover what correlations exist for the group studied and to test hypotheses concerning hypsodonty and mode of life.[21] Habitats considered were open (savanna, deserts), mixed (wooded savanna, brush) and closed (riverine and forest). Diets considered were grazing, mixed grazing and browsing, browsing, omnivory and special niches, such as treetop browsing. Measurements included the Hypsodonty Index (HI) and Muzzle Width (MZW). The results showed that, except for the "high-level" browsers, hypsodonty is correlated to "open and mixed habitats." The HI was not precise enough to discriminate between open and mixed. However, high MZW is correlated to grazing in the open category, although some forest species also have wide muzzles. Grazers therefore are distinguished by a combination of high-crowned cheek teeth and wide muzzles, both of which are possessed by E. Life in the open is implied. Cursorial distal limbs The preponderance of evidence is that E., as far as is known now, was a grassland grazer. It is in this context that Deng and Zheng, experts in the few surviving leg bones, conclude, concerning the morphology of the legs:[22] "Combined with hypsodont cheek teeth with much cement and strong enamel plication, the slender distal limb bones of E. caucasicum indicate that it is cursorial and dwells in an open steppe as a typical grazer." By distal they mean "furthest outward;" that is, the extremities. Cursorial animals are unequivocally "runners" although the authors did not examine what sort of gait scientifically should be proposed as "running." They selected caucasicum for study because of the availability of a few dozen limb bone fragments from Nihewan, China. These made possible a selective comparison with the fewer bones remaining of other fossil rhinocerotids. In comparison with them, the long legs of E. are the most derived; that is, the others did not have the same cursorial capabilities. The authors approach but do not solve the problem of how to reconcile the weight with the supposed mobility. They say elsewhere in the article that the legs of caucasicum are to be distinguished from those of other fossil Rhinocerotids at Nihewan by their "enormous size."[23] The White Rhinoceros at an estimated weight of 2.5 ton has been photographed galloping at a speed of about 30 kilometres per hour (19 mph). In a gallop, all feet are off the ground ("ballistic phase") twice a cycle, a feat that elephants, at 2.5-11 ton, cannot perform. They can walk up to 20 kilometres per hour (12 mph); however, their straight, relatively inflexible legs are those of striders, not the bent and spring-like legs of gallopers, which utilize haunches, ankle mobility and knee flexion to spring off the ground on alternative legs of a pair. E. legs are sufficiently like those of the White Rhino to hypothesize a similar gait even though E. weighed 4.5-5 ton.[24] Ceratomorphology The Ceratomorpha are so-called because their families, such as the Rhinocerotidae, of which E. is undisputably one, are characterized by the presence of hooves, or horns and hooves, made of keratin, the same substance of which hair is made. These keratin structures appear to have formed in the Mesozoic, a remnant in humans being the nails. A keratin horn is to be distinguished from a bone horn and a tusk. Bone forms the base of most horns but in some cases the horn is entirely of bone. A tusk is a modified canine or incisor tooth. Rhinocerotidae have had tusks, but not E. Two open questions are whether they were horned or hornless, hairy or hairless. Most Rhinocerotidae have and have had horns, but there are some instances of hornlessness, and most are or were hairy, such as the Wooly Rhinoceros, but no instances of hair or horn have yet been found for E. Only circumstantial evidence of them exists. The main evidence suggestive of a horn on E. is a frontal protuberance, which struck the attention of the late 19th century paleontologists and was immediately interpreted as the bony basis for a horn by most investigators from that time forward. A skull of E. sibiricum from the Volga region (cast shown in this article's lead picture) described by Alexander Brandt in the Russian journal, Niwa, and reported in Nature in 1878 offers the following description of the protuberance: hemispherical, 5 inches (13 cm) deep, furrowed surface, circumference of 3 feet (0.91 m). The furrows are interpreted as the seats of blood vessels for the tissues that generated the horn:[25] "The whole analogy with the rhinoceros points with the greatest certainty to the previous existence of a horn, which, to judge from the size of the blood vessels once encircling the base, must have possessed enormous dimensions." Brandt was already familiar with the legend of a unicorn among the Tatars of Siberia with a horn so large it required a sledge for transport, and made the connection in interpreting the bump as the base of a horn. He also interpreted the rostrum of the upper mandible as the basis of a nasal horn, a hypothesis now rejected in favor of the cropping labia. In any case the non-circular base indicates a section through the horn would not have been circular. This possibility is supported by another fossil with a non-circular partially healed puncture wound in the base, chiefly interpreted as the result of dueling other males with the horn.[14] The ungulates typically combine keratin and bone in various structures. If horns are keratinous, they have a bone core. Rhinocerotids horns, however, are uniquely derived. Hieronymus, an expert in Rhinoceros dermatology, says:[26] "...extant rhinocerotids are unique in possessing a massive entirely keratinous horn that approximates the functions of keratin-and-bone horns such as those of bovid artiodactyls ...." He defines rhinocerotid horns as: "...cylindrical blocks of constantly growing cornified papillary epidermis." This tissue is "strikingly convergent" with other "cornified epidermis" in horses, cetaceans, artiodactyls and birds. The horn is not attached to the bone of the boss but grows from the surface of a dense dermal tissue. The top layer keratinizes itself to form tubules about 1-2 millimeter high, the cells of which then die. The next layer forms below it.[27] As the layers age the horn loses diameter by degradation of the keratin due to ultraviolet light, desiccation and mechanical wear from contact with objects and agonistic behavior.[28] However, melanin and calcium deposits in the center harden the keratin there, causing differential wear and shaping of the horn.[29] The dermis generating the horn is anchored to the boss by interpenetration between rugosities – various irregularities of bone, which it creates by deposition.[30] This tissue is a specialization of dermal armor, which, whenever it attaches to bone, deposits the rugosities to strengthen the attachment. The author states that rugosity is "a bony signature of dermal armor."[31] Cranial rugisoty is an indication, but not a sure sign, of a horn. If, on the other hand, an annular (ring-shaped) pattern is visible in the rugosity, it is due to "stress concentration at the edges of horns" and is "the signature for epidermal horns." Hieronymus found annular rugosities in all living and some fossil Rhinocerotidae. The rings had previously been noted on additional fossils. To date E. has not been examined for rings under lighting designed to show them up; however, based on the observations of other paleontologists, the author says "squamosal rugosity" is the "most pronounced cranial rugosity in the elasmotherine lineage."[32] This fact suggests an especially firm attachment was required, which, combined with the extraordinarily large hump of muscle for managing the head, suggest a large and heavy horn. In the early 19th century the state of the fossils had not yet revealed the presence of a horn. By 1910 and for all the time since then the paleontologists have not ventured a mathematical estimate but have preferred to refer to the horn as immense, enormous, great or huge. As the size and shape of the horn depended on the concentration of melanin and calcium, and no known indicator of those remains, any further estimate of horn morphology is purely speculative. In 1968 Björn Kurtén's The Age of Dinosaurs pronounced, without a clue of the reasoning followed, that E. "carried a single frontal horn two meters in length."[33] This figure is often repeated in non-technical books and articles, as it is the only one available. The number is speculative, but coincidentally it happens to be the shoulder height of E. sibiricum. If in fact E. swept snow with the horn, or excavated holes in the terrain with it, the length can hardly have been less to reach the ground. Taxonomy The genus is known from hundreds of find sites, mainly of cranial fragments and teeth, but in some cases nearly complete skeletons of post-cranial bones, scattered over Eurasia from Eastern Europe to China. For example, Kazakhstan alone has 30 sites of E. sibiricum.[34] Dozens of crania have been reconstructed and given archaeological identifiers. The division into species is based mainly on the fine distinctions of the teeth and jaws and the shape of the skull.[35] The finds can be dated only by context. Classification within the rhinoceros family The classification of all the species and genera of the Rhinoceros family below the level of Rhinoceratidae has changed more rapidly in the first decade of the 21st century due to first the continued discovery of fossils representing new species and genera from China and Mongolia and second the number of points of morphology to be compared in computerized phylogenetic analysis. Two branches are generally recognized, that leading to the smaller Rhinoceroses and the Pliocene/Pleistocene branch of the larger elasmotheres. Both began from small species in the steppes of the Far East, but those in turn are comparable to North American species. Whether the branch point is considered to be at the subfamily level (Elasmotheriinae versus Rhinocerotinae) or at the tribe level (Elasmotheriini versus Rhinocerotini, the currently accepted version) or whether the elasmotheres existed at the subtribe level (Elasmotheriina, a discontinued taxon) is a matter of the status quo in publications, fossils and software programs. In 2002 P. O. Antoine performed a cladistic analysis using 282 "cranial, dental and postcranial characters"[36] of 28 "terminal taxa"[37] of "elasmotheriines from China and Mongolia" and four outgroups. He found that the elasmotheres were a monophyletic group. He says: "The main characters of derived elasmotheres (huge size, frontal horn, ossified nasal septum, enamel folding, hypsodonty, loss of anterior dentition, lengthening of the molar series ...) are absent in Middle Miocene Elasmotheriina from China and Mongolia. Most of these features appear somewhat later, during the Late Miocene or the Pliocene, in Parelasmotherium, Sinotherium and then Elasmotherium. Antoine was following the now discontinued practice of considering the Miocene ancestral species as elasmotheres. They were not, however, distinguished by the features of an elasmothere. The original ancestors were "minute brachydont animals." Elasmotherium was a "mammoth-sized hypsodont." In his view, "Such cranio-dental evolution demonstrates unequivocally the increasing proportion of grass-eating in the elasmotherine diet throughout the Middle Miocene." V.A. Terjaev in 1948 had a different view of the cause of the hypsodonty, maintaining that it was caused by the heavier grains in soil on plants pulled from moister environments, and that consequently Elasmotherians lived in "riparian biotopes."[38] Noskova points out that Elasmotherium's diet was comparable to that of the concurrent Archidiskon, which ranged over both steppe and riverland. Species Elasmotherium is believed to have descended from the Late Pliocene genus of Central Asia, Sinotherium. Elasmotherium is thought to be the most derived genus of elasmothere, with E. caucasicum in turn being more derived than E. sibiricum.[39] The two Chinese fossils, formerly considered distinct species, E. inexpectatum and E. peii, defined by Chow in 1958, have been sunk into E. caucasicum.[40] They were found in northern China from the Early Pleistocene Nihewan Faunal Assemblage (from the same valley as nearby Nihewan in Shanxi Province) and were extinct at approximately 1.6 Ma.[38] E. caucasicum, defined by Borissjak in 1914, flourished in the Black Sea region, as a member of the Early Pleistocene Tamanian Faunal Unit (1.1–0.8 Mya, Taman Peninsula). However, an elasmotherian species turned up in the preceding Khaprovian or Khaprov Faunal Complex, which was at first taken to be caucasicum,[41] and then on the basis of the dentition was redefined as a new species, E. chaprovicum (Shvyreva, 2004), named after the Khaprov Faunal Complex.[35] The Khaprov is in the MIddle Villafranchian, MN17, which spans the Piacenzian of the Late Pliocene and the Gelasian of the Early Pleistocene of Northern Caucasus, Moldova and Asia and has been dated to 2.6–2.2 Mya.[42] E. sibiricum, described by Johann Fischer von Waldheim in 1808 and chronologically the latest species of the sequence, coming from E. caucasicum in the Middle Pleistocene, ranged from southwestern Russia to western Siberia and southward into Ukraine and Moldova. It appears in the Middle Pleistocene Khazar Faunal Complex of the Sea of Azov region, which has "no exact stratigraphic situation."[43] The end of the elasmotheres is questionable; new evidence continues to turn up. The latest is from two caves in southern Siberia, a sibericum tooth in Smelovskaya Cave 11[44] and remains from Batpak,[45] both associated with Middle Pleistocene relict species, including large herbivores and predators. They must have been dragged into the caves by some predator, perhaps even modern man. The elasmothere tooth and one of a Cave Hyena from Smelovskaya were carbon dated to slightly greater than 50,000 BP in the Late Pleistocene.[46] A recent study of relict Late Pleistocene remains in the Beringia region (Alaska, Eastern Siberia) identified a pattern of "imbedded micrometeorites" consistent with an extinction event similar to the Tunguska event of 1908, only carbon-dated to 37,000 BP. A Siberian Elasmotherium skull in a museum was found to have this pattern.[47] Evolution The evolution of Rhinocerotidae follows a very similar pattern to the evolution of the Equidae, both being facets of the evolution of mammalian herbivores during the Cenozoic. The overall pattern is based on the evolutionary development of grass and grassland, and further the changing distribution of C3 and C4 types of plant metabolism, so-called because they generate a molecule containing 3 or 4 Carbon atoms by different metabolic pathways (sequences of chemical reactions). Evolution of Rhinocerotidae The Rhinocerotoids of the early Eocene and subsequently the Hyracodontidae and Amynodontidae show no sign of dermal armor or horns. The oldest known genus of Rhinocerotids, or true Rhonoceroses, is Teletaceras from the Middle Eocene of North America and the Late Eocene of Asia. Due to the timing of bridges, it does not appear in South America.[48] It features the first known Rhinocerotid tusks, a derived feature, which are still extant in three species and are "the primary offensive weapons" of those species. They appear to have been sexually dimorphic features convergent with the development of tusks in pigs, hippopotamoses and elephant seals.[49] The first dermal armor is indicated by cranial rugosity on Trigonias osborni, Late Eocene, 42-32 Mya. It appears more developed in Subhyracodon, early Oligocene, 33 Mya, both of North America.[50] There appears to have been a correlation between the tusks and the armor later (4 Mya later) defending against them.[51] Having begun to diversify (increase in species and genera) in America the Rhinocerotids spread rapidly over Beringia when that bridge appeared in the Early Oligocene to Eurasia, where subsequently they became highly diverse. The family reached a maximum floruit in the Miocene. At that time they entered Africa, but never were very diverse there. In the Pliocene they declined, disappearing from America. The Pleistocene brought extinction from Eurasia but African and Southeast Asian species survived.[48] Elasmotherium appeared in the Late Pliocene, apparently deriving from the Miocene—Pliocene Sinotherium, sometimes informally aggregated with it. Sinotherium in turn appeared to be related to Iranotherium; that is, Elasmotherium was believed to have been an iranotheriine. A 1995 cladistic analysis by Cerdeño asserted that Elasmotherium was most closely related to Coelodonta, the "Wooly Rhinoceros," whose soft-tissue corpus has been found in a few locations. By implication, Sinotherium was not an iranothere.[52] However, Antoine's 2002 analysis reaffirmed that the elasmotheriine line was iranotheriine. Certainty concerning the exact line of descent remains elusive; fossils and techniques of the future will perhaps resolve it. Evolution of grassland
The grass of grasslands belongs predominantly to the Poaceae family, also called the "true grasses." Species can be divided into "warm season grasses," which utilize C4, and "cool season grasses," which utilize C3. The evolution of grasses preceded the evolution of grasslands, which were created by the spreading of grasses from the forest onto the plains in the Miocene. These initial grasslands were C3, which happen to be more nutritious than C4 species.[53] The evolution of Rhinocerotidae and Equidae in the Miocene grasslands proceeded along parallel lines from similar causes. Diminutive, cursorial, browsing, brachydont species expanded their ranges from the Oligocene forests into the nutritious C3 Miocene grasslands and there over the millions of years of the Miocene evolved diverse and flourishing populations on the ample food supply, gradually increasing in size.[54] Some of these species exploited the grazing niche. Hispanotherium, Chilotherium and Teleoceras "show a clear tendency to hypsodonty, and a grazing feeding adaptation has usually been attributed to them." By the late Miocene, C4 metabolism was well established on the plains. In place of the prehistoric C3 grasslands (whuch are no longer extant) was the savanna, today's subtropical mixture of grasses and sedges. The deciding factors in this turn of evolution were a documented increase in temperature and aridity over the period. Faced with scant nutrition from browsing, the smaller Rhinocerotidae and Equidae declined in diversity, the gate in that direction, so to speak, now being closed.[55] The evolutionary response in the Pliocene was in the direction of bulk-feeding. Massive grazers, such as Elasmotherium, maximally hypsodont, moved across vast ranges consuming C4 grass in bulk. While the horses and other more nearly modern grazers could rely on galloping for mobility in any circumstance, there is a question as to what degree the brachypodial giants can be said to be "cursorial." Elephants, for example, must keep one foot on the ground, where true gallopers are entirely off it for brief intervals. The horns and impenetrable body armor insured that the giants would not be required to gallop away for any predator. At least until the arrival of man, they were, as the legends relate, lords of the savanna. Possible historical witnesses Legends and names referring to a special type of unicorn or mysterious beast with one horn, as opposed to the delicate imaginary unicorns of Europe based on narwhale horns, are scattered across the former range of Elasmotherium from China to Eastern Europe. They have been noted there since the first known literature of the Middle Ages. Eastern Eurasia The Chinese k'i-lin, referring to some sort of beast, was loaned in various forms into the Turkic and Mongolic languages and lore.[56] The writers of works in those languages have not known how to depict these strange beasts. A common theme is that they have a single horn, are dark and are huge. For example, a book of divination in the Old Turkic language uses kälän käyik, "tawny beast," (the kilin is altered to kälän by vowel harmony). The Han I Araha Manju Gisun I Buleku Bithe, "Mirror of the Manchu Language," dating from the 18th century, defines the kilin as[57] "...a quadruped with the body of a deer, the tail of a cow, the head of a sheep, the limbs of a horse, the hooves of a cow, and a big horn. His body is of five colors, his height a dozen feet. Of a gentle nature, he does not crush the worms or beak the plants." The Chinese had other unicorns, such as the Xie Zhi, but the latter was less like a real animal and more a symbolic figure. It had the ability to discriminate right from wrong and therefore a representaton of it was placed in the courtrooms of the Han Dynasty and, in the 3rd and 4th centuries, on the caps of judges. It looks like a seated or standing lion with a horn rising from the top of the head. In contrast to this mythical beast, the k'i-lin is presented as an animal actually caught by hunting parties of the past. The Shiji ("Historical Records") of Sima Qian in the 2nd century mentions the capture of a "deer-like" animal with one horn in 122 BC. The Chunqin, an encyclopedia of the 3rd century BC, states that a unicorn was captured in 481 BC, but without any description. Chinese representations of unicorns vary quite a lot, but an engraving on a bronze vessel of the Warring States Period, shows an animal very like the cave paintngs supposed to be of Elasmotherium: head down for grazing, horn protruding horizontally from the forehead, neck and shoulders humped.[58] In the northeast of the range, in 1866 Vasily Radlov reported a legend among the Yakuts of Siberia of a "huge black bull" killed by spear, who had "a single horn" so large it required a sledge for transportation. In 1878 A. F. Brandt suggested the beast was an Elasmotherium.[59] Western Eurasia From medieval northern Russia, probably Veliky Novgorod, a collection of ballads has survived, combined into a spiritual theme deriving ultimately from Zoroastrianism, but with Christian overtones, called the Stikh o Golubinioi knige, or in another version the Golubiniai kniga, "The Book of the Dove."[61] The "dove," or golub of the Christian work is probably an alteration of glub, "depth," so that the reconstructed name, *Glubinnaia kniga, is on the same theme as the Zend Avesta and similar works in Iran, the struggle between cosmic good and evil.[62] The force of pravda, or "rightness," is symbolized by a beast called mainly the Indrik, but also Indra, Beloiandrikh, Kondryk, Edinrog and Edinor.[63] The name is a calque: Greek mono-keros ("single-horn") > ino-rog (in, "one"; rogom, "horn") > edinrog > Indrik.[64] As a unicorn struggling with a lion representing krivda, "lies," the Indrik has the body of a goat or a horse in representations. An Indrik composite from all the variants of the ballads depicts the Indrik as dwelling in the Holy Mountain, which it consumes for sustenance. It is the mother or the father of all animals. It wanders unseen on the plain by day. When other beasts encounter it they must do obeisance. The Indrik saves the world from draught by digging springs of pure water. By night it wanders in the subterranean world, forging a path with its horn. There it purifies all the waters and releases them from any blockages. The righteous but fantastic unicorn appears also in the Bundahišn or Zand Āgāhīh, a Middle Persian Zoroastrian composition concerning the "deep and miraculous utterances" about the creation, the archetype of the *Gubinnaia kniga, "Book of the Deep." Its unicorn is an ass of 3 feet, 6 eyes, 9 mouths, 2 ears, 1 horn, a dark blue head and a white body, which is a symbolic rather than a real entity. It stands in the sea of Fraxwkand (wherever that is) and purifies the waters by urinating in them so that life may continue.[65] The Russian concept of a unicorn was apparently based on a different experience. In fact the Arabo-Persian word for unicorn in the region was karkadann, which extended also to India, and was loaned into the Turkmen language as kergeden and the Ottoman Turkish language as gergeden, among others. The word had an equivocal meaning: unicorn and rhinoceros.[66] This rhinoceros has capabilities and habits not possessed by the Indian rhinoceros: it indulges in mortal combat with elephants, which it kills by goring underneath with its horn. The symbol of evil is thus transferred to the elephant in that milieu; however, the two conduct their combats over grasslands never populated by either in historical times. The root of the word is Sanskrit Khadga-, Khadga dhenu-, "rhinoceros." In Christianity the single horn becomes a symbol of monotheism. Science writer and cryptozoologist Willy Ley theorised that these legends have simply been passed down since a hunting association with the beast in prehistoric times, and that the animals did not actually survive into recorded history.[67] One argument for survival is the testimony by the medieval traveller, Ibn Fadlan.[68] His account states:[69] "Near this river (the Volga) is a vast wilderness wherein they say is an animal that is less than a camel and more like a bull in size. Its head is like the head of a camel, and its tail is like the tail of a bull, while its body is like the body of a mule, and its hooves are like the cloven hooves of a bull. In the center of its head, it has a thick round horn, which as it rises from the head of the animal gets to be thinner until it becomes like the point of a lance. The length of some of these horns is from three to five cubits, and there are those that may attain to a greater or lesser length. The animal grazes on the leaves of trees, which are quite green. When it sees a horseman, it makes straight for him, and if he happens to have under him a fast horse, he is rendered safe from it with some effort. If it overtakes him, it removes him from the back of his horse with its horn, hurls him into the air, and then catches him with its horn. It continues in this manner until it kills him. It does not bother the horse in any form or manner. They seek out this animal in the forests in order to kill it. They do that by climbing the tall trees among which it is found, and with this object in mind, they assemble a number of archers with poisoned arrows. When it stands in their midst, they shoot at it until it is severely wounded and killed by them." "I saw in the king's house three large bowls which looked like [they were made of] the onyx of Yemen. The king informed me that it was made from the base of the horn of the animal. Some of the people of the country told me that it was a rhinoceros."
Notes ^ "Elasmotherium". PaleoBiology Database: Basic info. National Center for Ecological Analysis and Synthesis. 2000-. Retrieved 23 March 2011. References Antoine, Pierre-Olivier (March 2003). "Middle Miocene elasmotheriine Rhinocerotidae from China and Mongolia: taxonomic revision and phylogenetic relationships". Zoologica Scripta (The Norwegian Academy of Science and Letters) 32 (2): 95–118. doi:10.1046/j.1463-6409.2003.00106.x. Titov, V.V. (2008) (in Russian with English translations). Late Pliocene large mammals from Northeastern Sea of Azov Region. Rostov-on-Don: SSC RAS Publishing. Source: Wikipedia , Wikispecies: All text is available under the terms of the GNU Free Documentation License |
|